 |
Dr
Michel CAST 65 Avenue G. Clemenceau 40100 DAX [email protected] |
Accueil
Heliconius
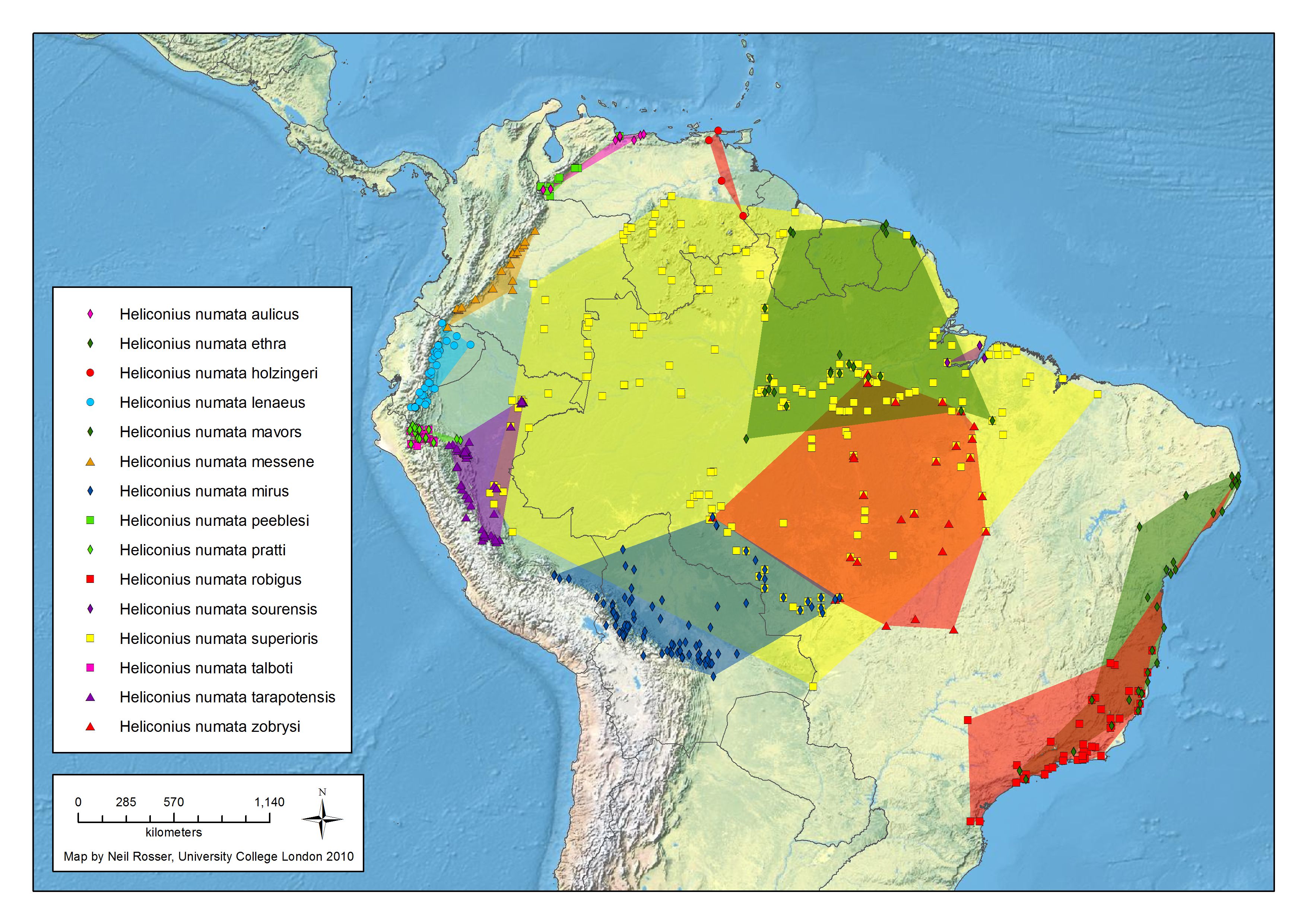
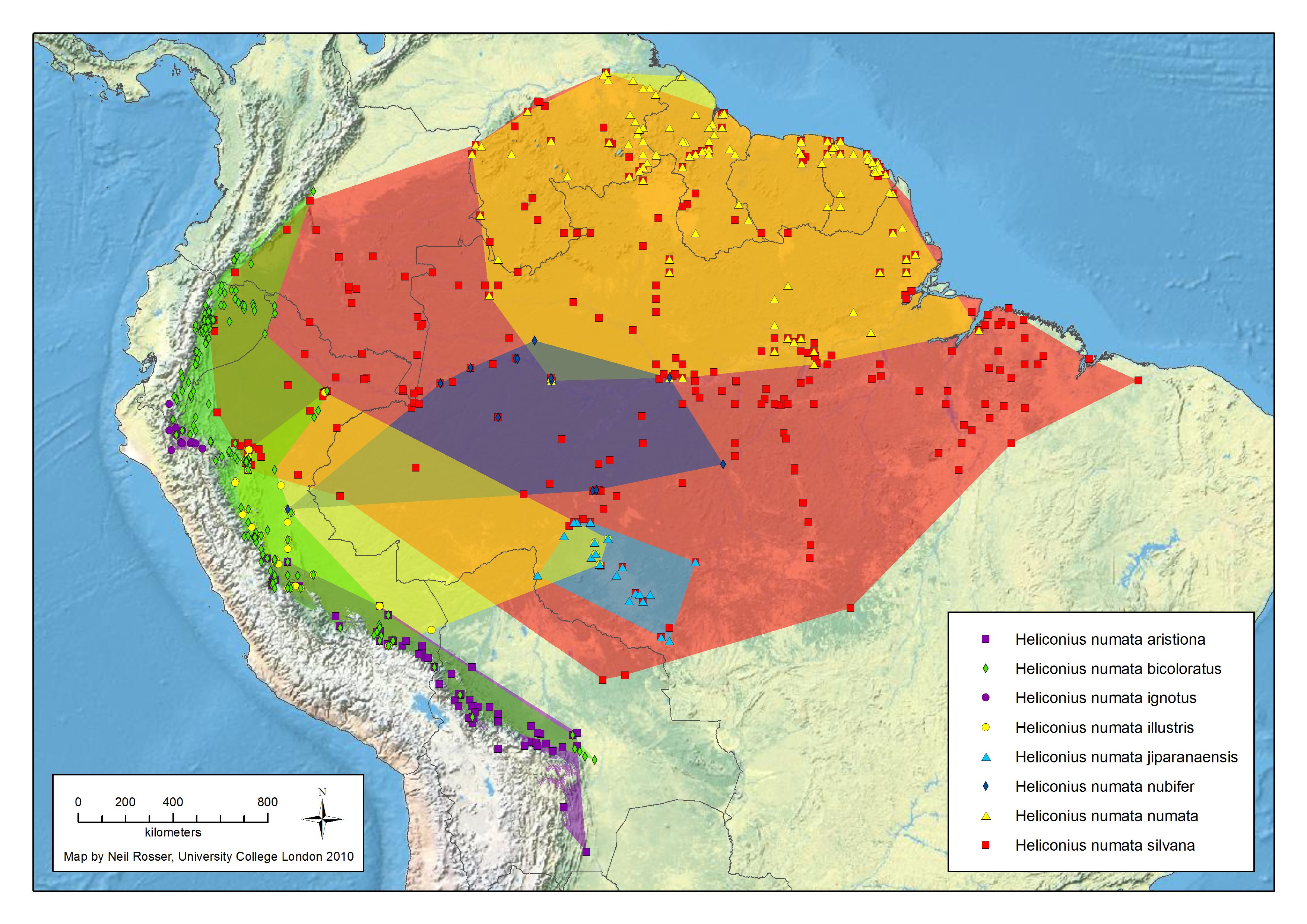
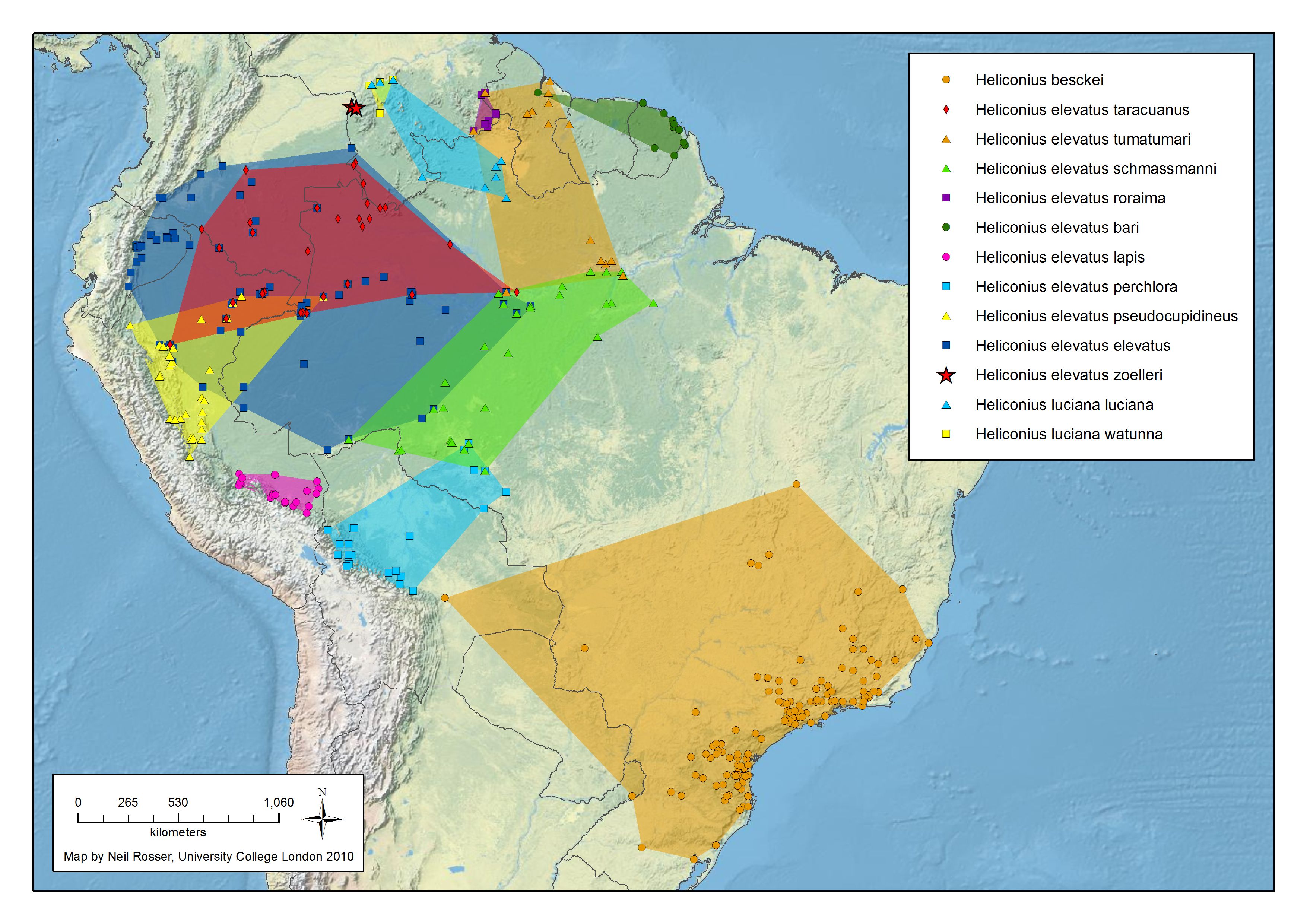
numata Cramer 1780
{kind=link}
{kind=link}
{kind=link}
{kind=link}
BROWN, K.
& BENSON, W. (1974) "Adaptive polymorphism associated with
multiple Mullerian mimicry in Heliconius
numata (Lepid. Nymph." Biotropica 6(4): 205-228
JORON, M. & Coll (2001) "Variable selection and the coexistence of multiple mimetic forms of the butterfly Heliconius numata". Evolutionary Ecology 13(7/8): 721-754 Lire PDF
JORON, M. & Coll (2006) "A conserved supergene locus controls colour pattern diversity in Heliconius butterflies". PLoS Biology 4(10): 1831-1840 Lire PDF
JONES, R., JORON, M. & Coll. (2012) "Evolution of a mimicry supergene from a multilocus architecture". Proceedings of the Royal Society of London (B) 279(1727): 316-325 (22 January; published online 15 June 2011) Lire PDF
JORON, M. & LE POUL, Y. & Coll. (2014) "Evolution of dominance mechanisms at a butterfly mimicry supergene" Nature Communications 5, Article number: 5644 Article
JORON, M. & Coll (2001) "Variable selection and the coexistence of multiple mimetic forms of the butterfly Heliconius numata". Evolutionary Ecology 13(7/8): 721-754 Lire PDF
JORON, M. & Coll (2006) "A conserved supergene locus controls colour pattern diversity in Heliconius butterflies". PLoS Biology 4(10): 1831-1840 Lire PDF
JONES, R., JORON, M. & Coll. (2012) "Evolution of a mimicry supergene from a multilocus architecture". Proceedings of the Royal Society of London (B) 279(1727): 316-325 (22 January; published online 15 June 2011) Lire PDF
JORON, M. & LE POUL, Y. & Coll. (2014) "Evolution of dominance mechanisms at a butterfly mimicry supergene" Nature Communications 5, Article number: 5644 Article
Génétique de la coloration alaire chez H. numata
H. numata possède une zone génétique appellée supergène qui commande différentes formes phénotypiques en fonction du mimétisme avec de nombreux Ithomidés. Au niveau de cette région chromosomiques, les scientifiques ont montré qu'un bloc d'une trentaine de gènes s'est trouvé immobilisé, au cours de l'évolution, par des phénomènes d'inversions (des régions d'ADN qui se retrouvent en position inversée chez différents individus), supprimant le processus naturel de mélange génétique entre générations. De cette manière, de nombreux gènes sont hérités en bloc et produisent des papillons d'apparence totalement différente, mais ressemblant à des espèces de Melinaea ou de Mechanitis. Cela permet de diminuer la possibilité d'avoir des individus aléatoires, éloignés de la ressemblance avec ces ithomidés. Cela permet, dans un espace géographique restreint, de "gérer" plusieurs motifs sans trop de formes intermédiaires.
D'après le CNRS, l'étude de JORON & Coll, "montre que 3 types chromosomiques sont en coexistence chez ces papillons, chaque type étant associé à une forme mimétique distincte. Ces 3 types chromosomiques sont distingués par des ordres de gènes différents, apparus par des événements d'inversion de certains segments d'ADN au cours de l'évolution. La variation de l'ordre des gènes réduit fortement les échanges génétiques entre chromosomes (recombinaison). Ceci explique que les 3 types chromosomiques ont un contenu ADN très divergent (sur le segment concerné) car ils ne sont jamais remaniés entre eux. Plusieurs gènes de coloration de l'aile présents dans cette région chromosomique se retrouvent ainsi couplés chez H. numata, grâce aux multiples inversions chromosomiques qui empêchent l'échange de gènes entre chromosomes. Ce couplage définit le supergène et explique son évolution, en fixant les segments d'ADN qui contrôlent les différentes formes mimétiques dans leur globalité, et empêchent leur remaniement."
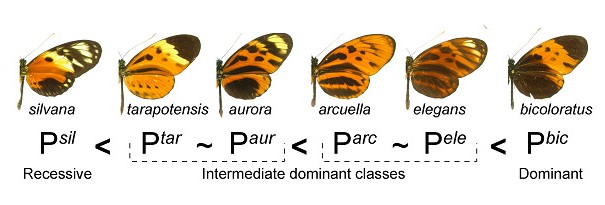
Les différentes formes ont génétiquement une importance de dominance différente. Par exemple, bicoloratus est dominante sur toutes les autres formes, alors que silvana est récessive.
Le supergène de H. numata:
L'essentiel du polymorphisme est géré par un supergène P avec jusqu'à 9 allèles (d'après Brown) qui est homologue avec le complêxe N-Yb-Sb de H. melpomene et du locus Cr de H. erato. Néanmoins, les gènes codant pour la synthèse des pigments ne sont pas liés à P.
| p(a) |
Augmente la
taille des points jaunes subapicaux |
| p(y) |
La bande
médiane jaune est située proximalement vers la cellule |
| p(d) |
La dague est
réduite ou absente |
| p(h) |
La bande
marginale noire des antérieures est réduite ou absente |
| p(c) |
La tache
médiane inférieure est absente |
| p(t) |
La tache en
virgule est large et triangulaire |
| p(s) |
La bande noire médiane des
postérieures est courbée vers l'extérieur et reliée à la bande
marginale noire par une aire triangulaire noire (cf silvana/superioris) |
| p(m) |
La bande noire médiane des
postérieures est très réduite et courbée vers l'extérieur (cf mirus/superioris) |
 |
 |
D'autres gènes sont impliqués qui modifient les patrons de colorations commandés par le supergène P:
Exemples d'après Brown:
| Phénotype |
Aspect
|
Exemples
|
| CH |
Suffusion
brune sur l'ensemble des ailes |
peeblesi,
leopardus |
| OR |
Une
suffusion orange remplace la couleur jaune |
silvana,
aristionia |
| AU |
Larges
points apicaux jaunes aux antérieures |
aurora |
| AP |
Pas de points apicaux jaunes aux antérieures | peeblesi,
messene, euphone |
| DH |
Suffusion
noire sur la plus grande partie des postérieures |
numata,
messene, aristionia |
| DP |
Une grande tache ronde
couvre la moitié des postérieures |
aurora,
euphrasius, confluens |
| DF |
Suffusion noire à la base
des antérieures |
messene,
aristionia |
| E |
La bande médiane noire des
postérieures séparées en plusieurs éléments |
euphone,
timaeus |
| EM |
Idem pour la bande
marginale noire des postérieures |
timaeus |
| LS |
Disparition presque
complète de la bande médiane des postérieures |
lenaeus |
| CL |
Bande médiane des
postérieures en forme de club de golf |
holzingeri |
D'autres d'après Jones, Joron & coll.:
Ces "petits" gènes, qui
interragissent avec P, semblent servir à affiner les patrons de
mimétisme, à les rendre plus ressemblants. Néanmoins, le léger
polymorphisme qu'ils induisent au sein d'une forme définie par le
supergène P a peut-être un coût en terme de prédation qui reste à
explorer.
On observe également qu'un certain nombre de gènes controlant le patron alaire sont associés au chromosome sexuel. On a donc un certain dimorphisme sexuel, mais a-t-il un impact sur une différence de mimétisme estre les mâles et les femelles?
On observe également qu'un certain nombre de gènes controlant le patron alaire sont associés au chromosome sexuel. On a donc un certain dimorphisme sexuel, mais a-t-il un impact sur une différence de mimétisme estre les mâles et les femelles?
Un article récent l'équipe de Mathieu JORON et Yann LE POUL a étudié la dominance au sein des allèles du supergène de H. numata. En croisant des individus de H. numata et en analysant les motifs colorés des descendants en découlant, les chercheurs ont découvert que la dominance entre allèles était déterminée par deux mécanismes distincts. L’un de ces processus semble être apparu lors de l’émergence de nouveaux types de motifs colorés, et fait en sorte que les allèles récents ont une dominance totale sur les allèles ancestraux.
Lire le résumé du CNRS et l'article.

La carte noir/orange décrit à chaque position de l'aile, la proportion d'individus hétérozygotes noir, orange, ou jaune (transcrit dans la couleur affichée) lorsque leur "parents" (homozygotes respectifs) sont noir ou orange. Une carte totalement noire indiquerait donc que à chaque fois que l'hétérozygote "à le choix" entre orange et noir, il "choisit" le noir, ce qui indique une dominance de la couleur noire sur le orange. Cette dominance dérivée de couleur n'a été observée qu'entre les allèles dérivés (première ligne) alors que les allèles ancestraux sont toujours récessifs envers les dérivés, quelque soir la coloration. (D'après Y. LE POUL com. pers.)